Looking at VA Mycorrhizae
"...it is usually impossible to tell whether or not a root system is mycorrhizal without staining and microscopic examination."
- Mycorrhizal Symbiosis, Smith and Read 1997
Mycorrhizal studies should be simple. VA mycorrhizae are abundant, free if one wants to take the time to collect field samples, quite reasonably priced if ordering pure cultures
from INVAM (the US organization for the distribution and maintenance of mycorrhizal cultures) and tolerant of a wide range of temperatures, soils, moisture levels, and plant hosts. In practice, though, mycorrhizae are
quite difficult to study, and development of simpler and more exact techniques has occupied much of the time and energy of the field.
There are several obstacles for researchers to overcome:
1) Mycorrhizae are obligate symbionts. Although a few species of ectomycorrhizae have been cultured in nutrient agar, I am not aware of any endomycorrhizae that have been
cultured without plant hosts.
2) VA mycorrhizae grow inside opaque plant roots, necessitating some technique for making roots transparent or looking through the roots. (Extraradical hyphae can be seen
with only the help of a light microscope.)
3) Soil is even more opaque than roots, making field studies of VA mycorrhizae by their very nature artificial. Most techniques for studying VA mycorrhizae in the field
involve construction of clear walls against which mycorrhizal roots grow, or the removal of mycorrhizae from soil.
4) 100% mycorrhizal infection is rare. Techniques for quantifying the amount of root area infected by mycorrhizae must overcome the difficulties of multiple entry points of
hyphae into a root and the variation, even within a single root, in the presence and abundance of vesicles, arbuscules, spores, and hyphae.
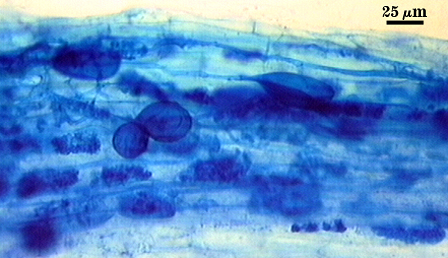 With these difficulties taken into account, it is not hard to realize why the staining technique of choice today was developed
almost 30 years ago. J.M. Phillips and D.S. Hayman (1970) published the trypan blue staining procedure that is still a standard in the
field. The basic technique involves collecting roots and cutting them into 1 cm segments. These root segments are cleared for varying
amounts of time (depending on plant species and root thickness) in 10% KOH solution, often at high temperatures. Cleared roots are
rinsed and placed in a weak trypan blue solution (0.05% to 0.1% wv dye concentrations are common), which stains mycorrhizae dark
blue while leaving the stele of some roots light blue and most root cells an even paler blue. Stained roots are destained in 10% lactic
acid, and stored in lactic acid solution or glycerol. Stained roots can be examined under low magnification (as low as 40x for some species; 100x or more for more careful
quantification and systematic studies) using a light microscope, and techniques for quantifying mycorrhizal infection abound.
With these difficulties taken into account, it is not hard to realize why the staining technique of choice today was developed
almost 30 years ago. J.M. Phillips and D.S. Hayman (1970) published the trypan blue staining procedure that is still a standard in the
field. The basic technique involves collecting roots and cutting them into 1 cm segments. These root segments are cleared for varying
amounts of time (depending on plant species and root thickness) in 10% KOH solution, often at high temperatures. Cleared roots are
rinsed and placed in a weak trypan blue solution (0.05% to 0.1% wv dye concentrations are common), which stains mycorrhizae dark
blue while leaving the stele of some roots light blue and most root cells an even paler blue. Stained roots are destained in 10% lactic
acid, and stored in lactic acid solution or glycerol. Stained roots can be examined under low magnification (as low as 40x for some species; 100x or more for more careful
quantification and systematic studies) using a light microscope, and techniques for quantifying mycorrhizal infection abound.
There are several problems with the trypan blue staining method. First, trypan blue is a carcinogen. The slightly less toxic dye chlorazole black E (CBE) has been suggested as a
possible alternative for root staining. The trypan blue technique does not distinguish live mycorrhizae from dead mycorrhizae, making it less useful for studies that relate the amount
of mycorrhizal infection to the impacts of that infection on plant hosts. Not only are dark blue mycorrhizae on a light blue background difficult to see, but trypan blue will
occasionally stain root cells, very thin root segments, the stele of the root, or non-mycorrhizal fungi the same color as the target mycorrhizae. And finally, mycorrhizae in roots
subjected to this staining technique are often, as one might predict, broken or altered by the clearing process.
To attempt to solve these problems with an otherwise effective (and gratifyingly simple) technique, and to take advantage of recently developed stains, several other staining
techniques are coming into use in the examination of VA mycorrhizae:
Acid fuchsin:
(from S. Dickson and S.E. Smith in Varma 1998)
Acid fuchsin, along with chlorazole black E (CBE), can be used in similar ways to trypan blue. Both of these stains show both live and dead mycorrhizae,
as well as staining some non-mycorrhizal fungi and plant structures. Acid fuchsin, which stains mycorrhizae pink, is often used in combination with vital
stains (especially nitroblue tetrazolium, NBT) to show the inactive mycorrhizal structures that are not detected with enzyme-specific stains.
Vital stains:
(from S. Gianinazzi and V. Gianinazzi-Pearson in Norris et al. 1994; S.Dickson and S.E. Smith in Varma 1998)
Vital stains separate active mycorrhizal structures from structures with no enzymatic activity. Nitroblue tetrazolium (NBT) is a commonly used vital stain that is coupled to the
activity of succinate dehydrogenase, an enzyme that is involved in the Kreb's cycle. Active mycorrhizal structures (those that contain
active succinate dehydrogenase) stain purple. As mentioned above, counterstaining with a non-vital stain such as acid-fuchsin allows the detection of both active and inactive
mycorrhizae.
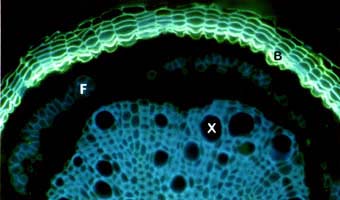
Fluorescein diacetate (FDA) is hydrolysed by
esterases in mycorrhizal cells, and causes those cells to become fluorescent. Active mycorrhizae glow green, while dead mycorrhizal structures are not stained. Unfortunately, FDA
also stains the cell walls of plant roots, and the fluorescence of these cell walls interferes with the observer's ability to measure mycorrhizal infection. FDA can only be used
accurately with root sections.
A more specific vital stain, alkaline phosphatase (ALP) stains only fungal structures. ALP-stained mycorrhizae are violet-black. There is conflicting evidence about ALP's usefulness
as a marker of mycorrhizal structures involved in phosphorus uptake. Tisserant et al. (1993 in Varma 1998) found that ALP activity in intraradical hyphae increased prior to a
growth response of the host plant. However, Larsen et al. (1996 in Varma 1998) found that mycorrhizae that had been treated with a fungicide showed reduced phosphorus
transport, but no reduced ALP activity. Regardless, ALP is a more specific stain for mycorrhizae than FDA and has been used with root segments as well as root sections.
Antibody staining:
(from S. Gianinazzi and V. Gianinazzi-Pearson in Norris et al. 1994)
Gold-labeled antibodies raised against proteins found only in mycorrhizae can be used to determine the internal and external mycorrhizal structures in root sections. Although
electron microscopy is probably the best way to examine root sections with this technique, light microscopy (with as little as x135 magnification) is also effective. Areas
containing the target of the gold-labeled antibodies appear dark and contrast sharply with the rest of the root section. Specific antibodies (e.g. for b1-protein; See
Gianinazzi-Person et al. 1991) seem to show that defense responses by plant cells to VA mycorrhizal infection are not transferred to nearby uninfected cells. Antibody staining
is one of the most promising techniques for investigating the differences between mycorrhizal structures, but, like several of the vital stains and electron microscopy techniques,
can only be used with root segments.
Auto-fluorescence:
(from Friese and Allen 1991)
VA mycorrhizae inside plant roots are difficult to see and to quantify, but arbuscules, conveniently, autofluoresce under ultraviolet light. As Gianinazzi and
Gianinazzi-Pearson (in Norris et al. 1994) point out, this is one of the few non-destructive ways to observe mycorrhizae. Live arbuscules glow yellow under violet-blue light,
allowing observation of mycorrhizal development over time. When paired with direct observation of external hyphae under a dissecting microscope, this technique can be
particularly effective and allow mapping of the spread of mycorrhizal infection. Unfortunately, both auto-fluorescent observation and direct observation of external hyphae
require the plants used to be grown against clear glass or plastic, a highly artificial situation that may very well change the architecture of mycorrhizal infection.