Biology Dept Kenyon College |
Limb Development |

|
Biology Dept Kenyon College |
Limb Development |
|
|
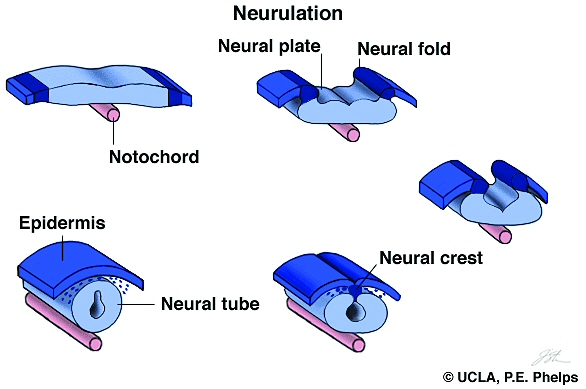
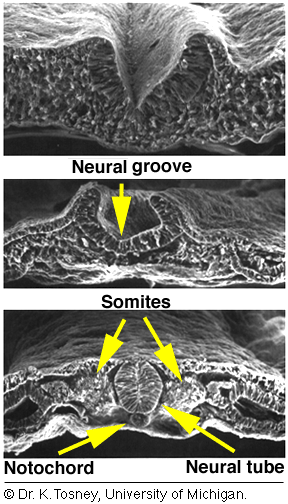
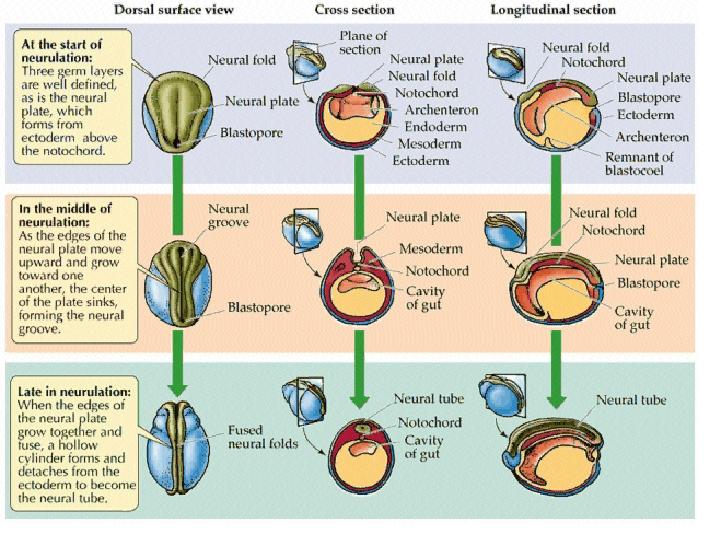
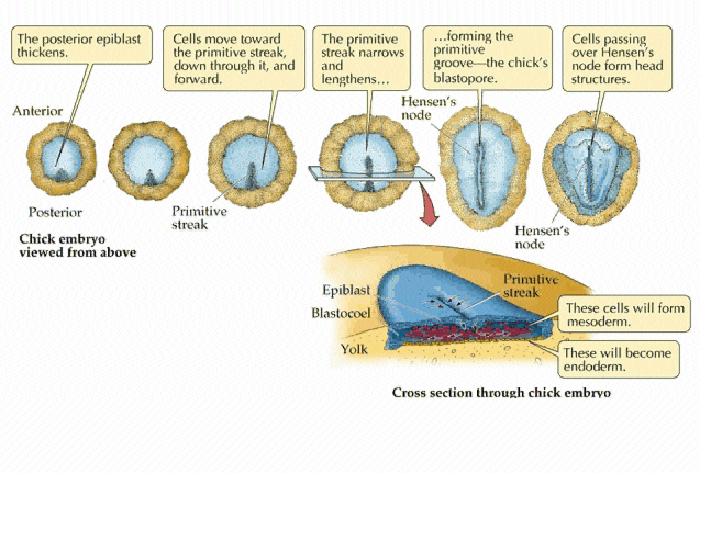
Neurulation Somites and Hox genes (below) Neurulation in vertebrates results in the formation of the neural tube, which gives rise to both the spinal cord and the brain. Neural crest cells are also created during neurulation. Neural crest cells migrate away from the neural tube and give rise to a variety of cell types, including pigment cells and neurons. Neurulation begins with the formation of a neural plate, a thickening of the ectoderm caused when cuboidal epithelial cells become columnar. Changes in cell shape and cell adhesion cause the edges of the plate fold and rise, meeting in the midline to form a tube. The cells at the tips of the neural folds come to lie between the neural tube and the overlying epidermis. These cells become the neural crest cells. Both epidermis and neural plate are capable of giving rise to neural crest cells. What regulates the proper location and formation of the neural tube? The notochord is necessary in order to induce neural plate formation. Below are scanning electron micrographs of a chick embryo during neurulation.
During neurulation, somites form in pairs flanking the neural tube. Somites are blocks of cells that form a segmental pattern in the vertebrate embryo. Somites produce cells that become vertebrae, ribs, muscles, and skin. The region where neural tube closure begins varies between different classes of vertebrates. In amphibians such as Xenopus, the neural tube closes almost simultaneously along its entire length. In birds, the neural tube closes in the anterior to posterior direction, as Hensen's node regresses. Mammalian neurulation is similar to that of birds, however the bulky anterior neural plate seems to resist closure - the middle of the tube closes first, followed by both ends. Watch this animation of mammalian neurulation!
This video of a living Xenopus (frog) embryo shows both gastrulation and neurulation. You should recognize the beginning of the film from our discussion of gastrulation. The open neural plate on the dorsal side has formed by the time the blastopore closes. The closure of the neural plate into a tube is accompanied by elongation of the embryo.
Another movie of Xenopus development is available here!
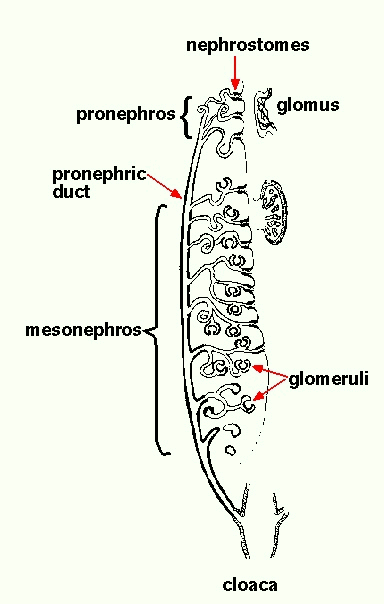
Animal development: Organogenesis Organogeneis is the period of animal development during which the embryo is becoming a fully functional organism capable of independent survivial. Organogenesis is the process by which specific organs and structures are formed, and involves both cell movements and cell differentiation. Organogenesis requires interactions between different tissues. These are often reciprocal interactions between epithelial sheets and mesenchymal cells. 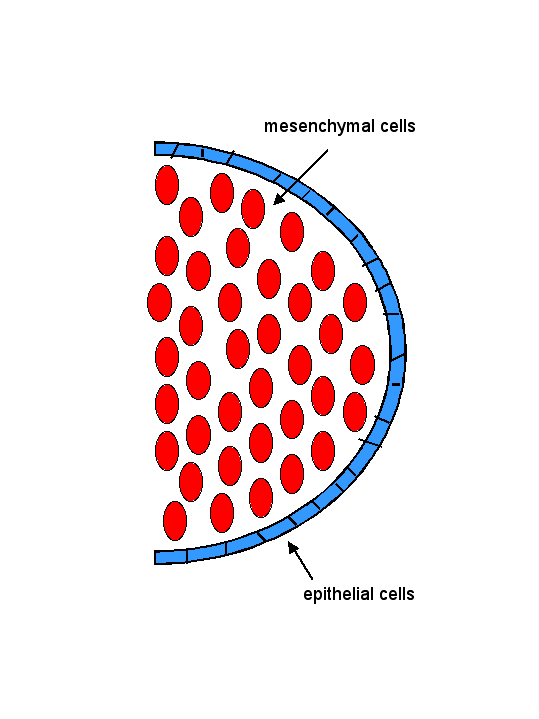 The study of organogenesis is important not only because of its relevance to understanding fundamental mechanisms of animal development, but also because it may lead to medical applications, such as the repair and replacement of tissues affected by genetic disorders, disease or injury. Kidney development There are three stages of mammalian kidney development: the formation of the pronephros, mesonephros, and metanephros (nephros = kidney; pro = before, meso = middle, meta = after). The metanephros is the permanent kidney found mammals (and in birds and reptiles), and forms at the region between the mesonephros and the cloaca (below).
The development
of the adult kidney (metanephros) provides a good example of reciprocal
epithelial-mesenchyme interactions. Mature (metanephric) kidneys form
from reciprocal inductions between the metanephric mesenchyme and the
(epithelial) ureteric buds. Metanephric
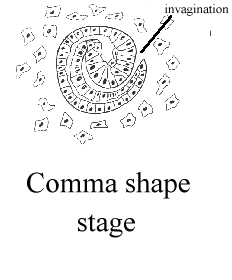
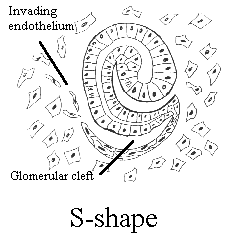
kidney development is a multistep process. 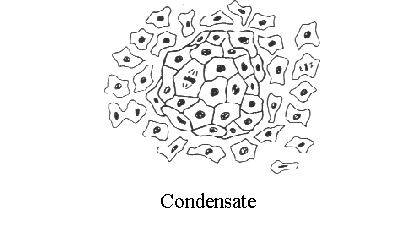 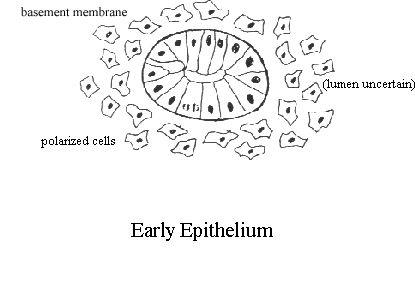 images from the Kidney Development Database 3. Each aggregate forms a nephron: first a comma shape is observed, and then the S-shaped tubule, which connects to the branched ureteric bud
What is the
experimental evidence for reciprocal induction?
Vertebrate limb development Vertebrate limbs
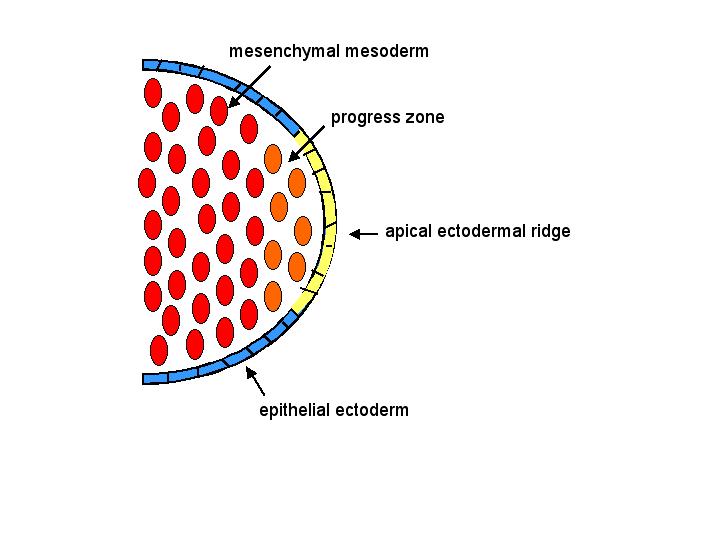
develop from limb buds. The vertebrate limb bud consists of a core of loose mesenchymal mesoderm covered by an epithelial ectodermal layer. Cells within
the progress zone rapidly divide,
and differentiation only occurs once cells have left the progress zone. Because of this process,
differentiation proceeds distally as the limb extends (that is, the
proximal end of the limb develops before the distal end). The apical ectodermal ridge at tip of limb bud
induces the formation of the progress zone.
Pattern formation organizes cell types into their proper locations based on positional information. Anterior-posterior patterning is regulated by the zone of polarizing activity, or ZPA. The current model is that proximal-distal pattern formation is regulated by the amount of time a cell spends in the progress zone. Dorsal-ventral patterning is controlled by the overlying ectoderm. What makes forelimbs
and hindlimbs different from one another? Pattern formation is
regulated by the same signals in both limbs, although these
signals are interpreted differently. Limb-specific transcription
factors have been identified, and by expressing these transcription
factors in the OTHER (wrong) limb, scientists have been able to observe
transformation of the hindlimb into the forelimb, and vice-versa.
from the Max Planck Society Thanks to David Marcey for construction of some of the images shown above |

{kind=link}