Immunogenicity of HBV: Even though both humoral and cell-mediated immunity (CMI) result from natural HBV infection, the presence of antibodies alone is enough to provide protection from infection or reinfection. The exact role of CTLs in protection or clearance of the HBV infection is not yet known. After natural infection, antibodies (anti-HBs) are detected against the HBsAg and viral core antigen (HBcAg, anti-HBc). There is a very clear role for anti-HBs in providing immunity and all vaccines used in humans have been designed to evoke this. In humans a level of 10 mIU/ml (milli-international units/ml) is considered sufficient to confer protection. The role for anti-HBc in providing immunity is less clear. It is known that HBc is highly immunogenic but the protective effect of the anti-HBc antibodies is still in question since chronic carriers generally have high titers of anti-HBc, but not anti-HBs (Davis et al., 1998).
Problems with current HBV vaccines: The
vaccines presently used to immunize patients against HBV are subunit vaccines
consisting of empty subviral particles composed solely of HBV envelope
protein. Initially these were obtained from the plasma of chronic carriers
of the virus, but their use was restricted by a limited supply of chronically
infected plasma and by concerns about their safety. Currently, the majority
of HBV vaccines contain recombinant HBsAg protein produced in stable transfected
yeast or hamster ovary cells (Jilg, 1998). Although
these recombinant protein vaccines are highly effective, they are expensive
to produce, which inhibits there use in the third world areas where they
are needed most. These vaccines also have disadvantages; multiple doses
are required that each dose must be kept cold, both of which further
increase their cost in areas where  refrigeration
is not common. Other shortcomings of the subunit vaccines include non-response
which occurs when there is no detectable level of anti-HBs in the patient
after vaccination, or hypo-response where there is less than the sufficent
level of anti-HBs required for protection despite repeat injections. Reseachers
have been working dilligently for the past few years to find an effective
yet inexpensive vaccine. It is not possible to use a whole killed or live
attenutaed vaccines due to the difficulty of growing HBV in culture and
also due to the undesirable potential for improper reactions in immunocompromised
patients. There is also a great need around the world for a therapeutic
vaccine which could be used instead of the current HBV treatments using
interferons, which are only at best partially effective, to treat chronically
infected individuals (Daivs, 1998).
refrigeration
is not common. Other shortcomings of the subunit vaccines include non-response
which occurs when there is no detectable level of anti-HBs in the patient
after vaccination, or hypo-response where there is less than the sufficent
level of anti-HBs required for protection despite repeat injections. Reseachers
have been working dilligently for the past few years to find an effective
yet inexpensive vaccine. It is not possible to use a whole killed or live
attenutaed vaccines due to the difficulty of growing HBV in culture and
also due to the undesirable potential for improper reactions in immunocompromised
patients. There is also a great need around the world for a therapeutic
vaccine which could be used instead of the current HBV treatments using
interferons, which are only at best partially effective, to treat chronically
infected individuals (Daivs, 1998).
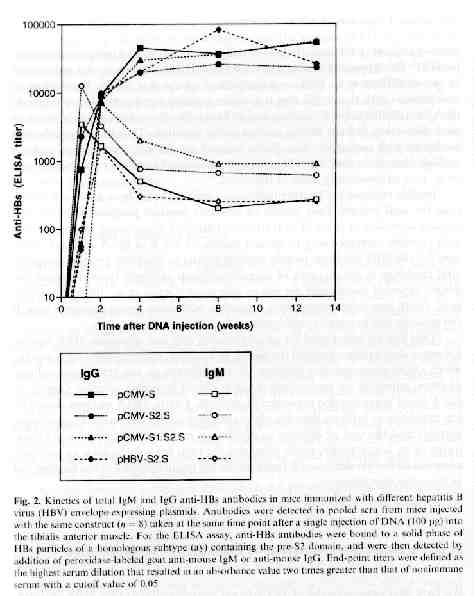
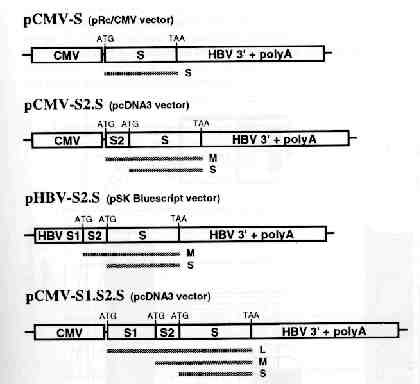
HBV DNA vaccine model: H.L. Davis et al. (1998) presented a model for a HBV DNA vaccine that used four vectors that expressed one or more of the HBV envelope proteins that are encoded by a single gene within the HBV genome. This gene is divided into S, pre-S2 and pre-S1 regions by three ATG start codons. Three of these four vectors used for the vaccine utilized the immediate early promoter of cytomegolovirus (CMV) to drive the expression of the S (pCMV-S) pre-S2 + S (pCMV-S2.S) or pre-S1 + pre-S2 + S (pCMV-S1.S2.S) envelope gene. The forth vector uses the endogenous HBV promoter within the pre-S1 domain to drive expression (pHBV-S2.S) (Davis et al., 1998). This model used again by H.L Davis, and then by M-L Michel et al. in 2001. In 2001, R. Schirmbeck et al. (2001) developed a modified version of this model, using the commercially available pCI expression vector instead of the pCMV as a promoter to drive the expression of soley the S vector (pCl-S). A similar model using pCI was also used by A. Thermet et al. in 2003. All of these five studies used intramuscular injection of the HBsAg-expression plasmids in murines used to model human response. R. Schirmbeck et al. also opted to use subcutaneous injections and DNA coated gold particles administered with a gene gun (Shirmbeck et al., 2001) .
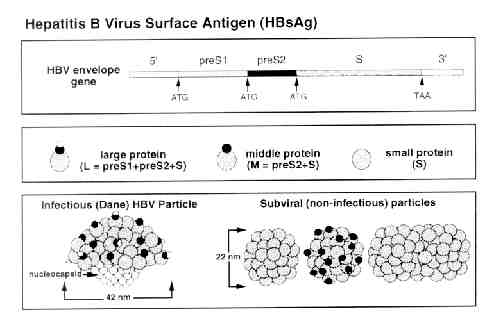
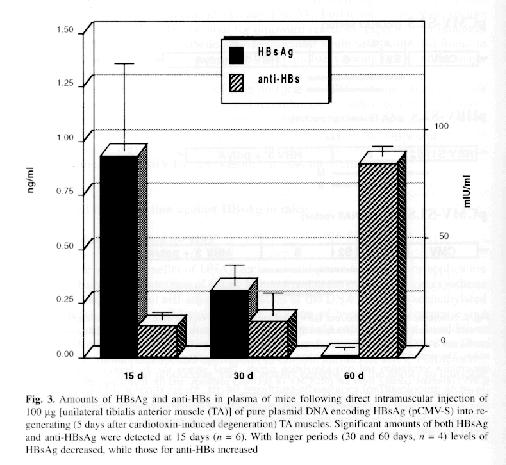
In vivo synthesis of HBsAg and anti-HBs: H.L. Davis et al. (1998) found that HBs-Ag expression muscle fibers retained a normal histological appearance 5 days after DNA injection, but were seen to breakdown at about 10 days and completely disappear after 30 days. They determined that the disappearance of these muscle fibers was probably due to attack by HBsAg specific CTL. Low levels of circulating antigen were detected early on in the study and undetectable amounts of HBsAg were found in the serum later, which reflects the neutralization of antigen by antibody as well as a decrease in the production of HBsAg with the destruction of the fibers (Davis et al., 1998). Similar findings were described again by Davis in her study of DNA vaccine immunization against HBV in animal models. R. Schirmbeck et al. found similar serum antibody levels, noting that the highest antibody production was found when just the HBsAg antigen were injected instead of the vectors encoding for HBsAg (Schirmbeck et al., 2001). A. Thermet et al. (2003) also recorded similar findings, describing a significant increase in anti HBsAg antibodies after the initial injection of the protein encoding vectors (Thermet et al., 2003).
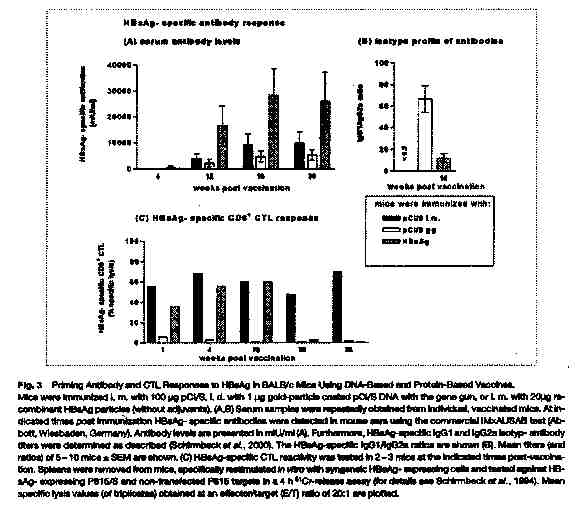
(Schirmbeck, S. et al.)
(Davis, H.L., et al.)
Humoral response: Immunization of murines by interamuscular injection of plasmid DNA that express HBV envelope proteins and HBsAg stimulate fast, strong and long lasting humoral immune responses. Antibodies, which are initially of the IgM then IgG isotype, are produced; they are able to recognize several of the B-cell epitopes on the S, preS2 or preS1 domains of the envelope. (Michel, et al., 2001). With S-encoding DNA, there is a predominance of group-specific antibodies, which are capable of providing protection against certain strains of HBV. After injection, the pre-S2 domain predominates, especially in the early period. Pre-S2 antibodies are believed to to be beneficial for prophylactic vaccination due to the fact that they are known to produce protective immunity in chimpanzees and other animal models (Davis et al., 1998). High titers of anti-HBs were found by M.L. Michel et al. (2001) by 4-8 weeks and persisted for at least 17 months after a single DNA injection. Antibody concentration was boosted ten-fold after the second injection of DNA, and somewhat less by injection of a recombinant HBsAg protein (Michel et al., 2001).
Cell mediated responses: DNA-based immunization using HBsAg plasmids result in strong CTL response. Murines injected with vectors expressing any of the three forms of the HBsAg protein produced high levels of CTL and CTL precursors. After prolonged stimulation with HbsAg-expression cells, spleen cells from DNA-immunized murines were capable of specific lysis of the present antigen, indicating high levels of CTL precursors. CTL activity developed between 3 and 6 days after DNA injection and reached maximal levels by day 12, expressing peak activity for months (Michel et al., 2001).
Potential for HBV DNA vaccines in humans: Results following injection of HBsAg-encoding plasmid DNA in murine models indicate that the DNA vaccine should be feasible for vaccination in humans against HBV. A DNA vaccine would be superior to recombinant HBsAg for prophylactic vaccination; this is especially true in cases of non- or hypo- responsiveness, or for the induction of a rapid humoral response in infants born to chronic carriers (Davis et al., 1998). There have been a few promising clinical trials which reported that DNA vaccines appear to be well tolerated in human hosts. Despite the induction of significant CTL responses, vaccination via needle or gene gun administration of DNA failed to induce antigen-specific antibodies. Increased efficiency could come from improvements in the tranfection or in the induction of better immune responses through the coexpression of cytokines or combinations with other non-DNA vaccines. However, due to the induction of strong CTL, DNA vaccines may be effective for the treatment of chronic HBV patients (Michel et al., 2001).