Biology Dept Kenyon College |
|

|
Biology Dept Kenyon College |
|
|
| Differential gene
expression and
development
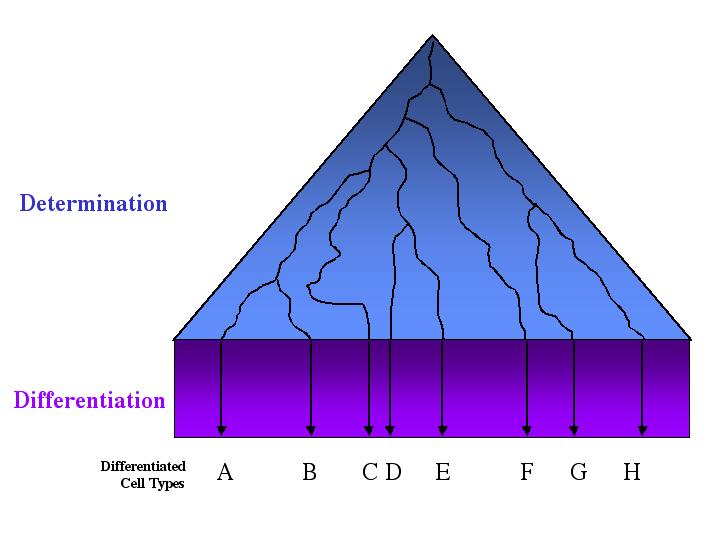
Mechanisms of cellular determination Induction Pattern formation Differential gene expression and development The fate of a cell describes what it will become in the course of normal development. The fate of a particular cell can be discovered by labelling that cell and observing what structures it becomes a part of. When the fate of all cells of an embryo has been discovered, we can build a fate map, which is a diagram of that organism at an early stage of development that indicates the fate of each cell or region at a later stage of development. The developmental potential, or potency, of a cell describes the range of different cell types it CAN become. The zygote and its very early descendents are totipotent - these cells have the potential to develop into a complete organism. Totipotency is common in plants, but is uncommon in animals after the 2-4 cell stage. As development proceeds, the developmental potential of individual cells decreases until their fate is determined. The determination of different cell types (cell fates) involves progressive restrictions in their developmental potentials. When a cell “chooses” a particular fate, it is said to be determined, although it still "looks" just like its undetermined neighbors. Determination implies a stable change - the fate of determined cells does not change. Differentiation
follows
determination, as the cell elaborates a cell-specific developmental
program.
Differentiation results in the presence of cell types that have
clear-cut
identities, such as muscle cells, nerve cells, and skin cells.
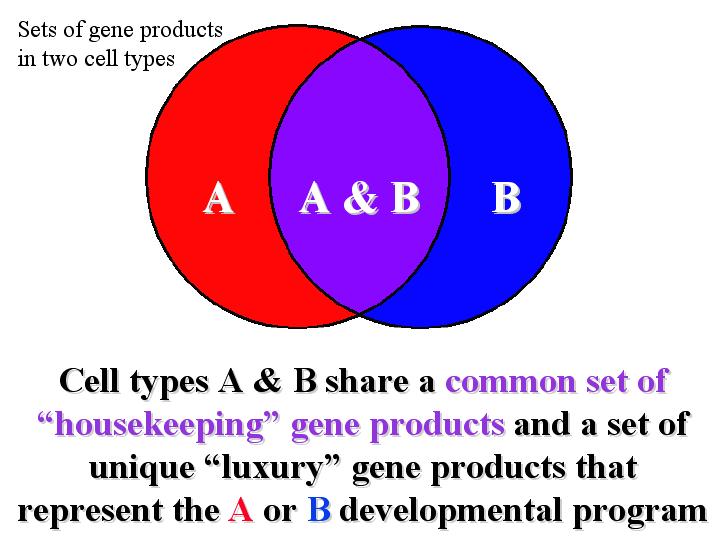
Differentiation results from
differential gene expression:
The specific components of a given cell provides its special
characteristics.
These components are either synthesized by proteins, or are themselves
proteins. By expressing different subsets of genes, two cells contain
different
subsets of gene products (proteins).
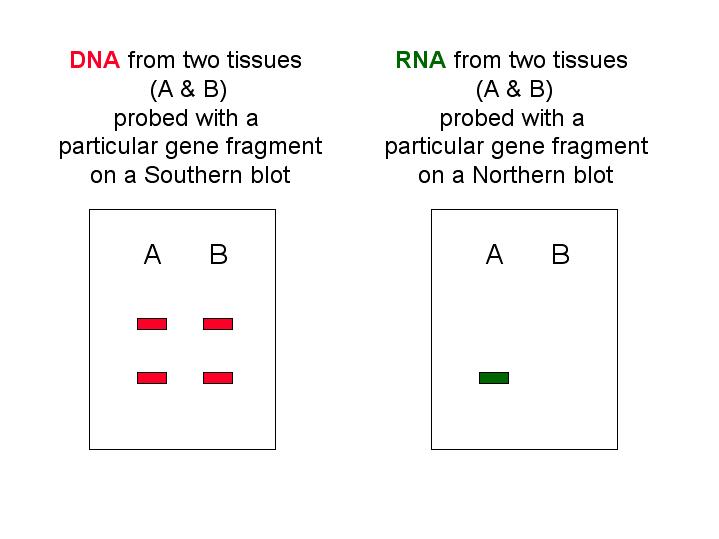
How can we observe that cells from two tissues express different genes? Below are two blots: the Southern blot shows that tissues A and B both contain a particular gene. However, the Northern blots shows that only tissue A contains RNA transcribed from that particular gene.
Differential gene expression is not a result of differential loss of the genetic material, DNA, except in the case of the immune system. That is, genetic information is not lost as cells become determined and begin to differentiate.
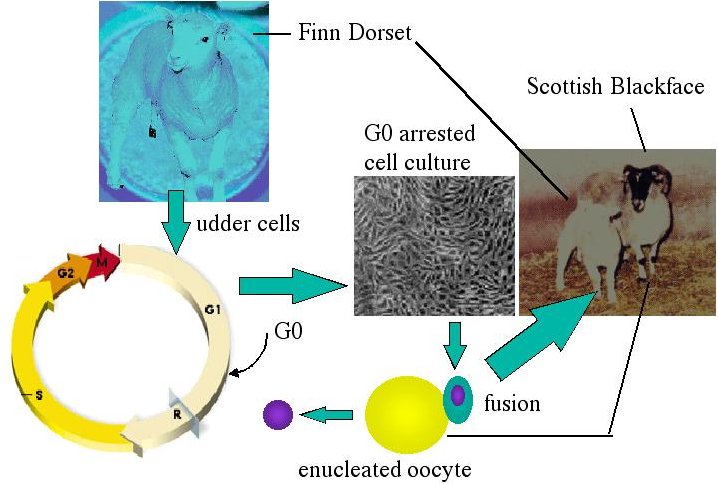
In order to clone Dolly, udder cells were removed from a Finn Dorset ewe and starved for one week to cause G0 arrest. Nuclei from arrested Finn Dorset udder cells were fused with enucleated eggs from a Scottish Blackface ewe, and then stimulated to re-enter the cell cycle. After a few rounds of cell division, the embryo was transplanted into a surrogate Scottish Blackface mother. The sheep that was born was genetically identical to the Finn Dorset ewe, which was the source of the nucleus.
Transplantation of imaginal discs in insects and the cloning of whole plants from individual cells strengthens the conclusion that genetic information is not lost as cells become determined and begin to differentiate. While differentiation results in specific cell types, morphogenesis is the process whereby the shape (morph) of the embryo is generated (genesis). Morphogenesis in both plants & animals involves regulated patterns of cell division and cell elongation that leads to changes in cell shape. Cell movement also plays a critical important role in animal morphogenesis. How do cells become different from their
parent cells?
How do two identical daughter cells become different from one another?
How might one daughter cell become a neuron, while the other daughter
cell
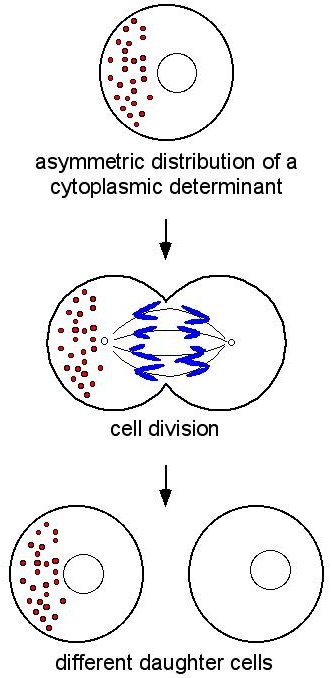
becomes a skin cell? In some cases, determination results from the asymmetric
segregation of cellular determinants. However, in most
cases,
determination is the result of inductive
signaling between cells.
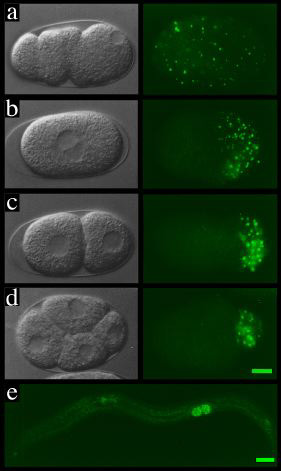
Movie of assymetric cell segregation Although there are many examples where the
asymmetric segregation of cellular determinants leads to differences
between daughter cells, more frequently we find that cells become
different from one another as a result of inductive signals coming either
from other cells or from their external environment. Although one of the classic models for signaling involves diffusion, there is new evidence that inductive signals may in fact be actively transported within and between cells, and that cellular projections may be involved in long distance communication between cells. How do organs develop in their proper positions? How do cells "know" where they are within a developing organism? Pattern formation concerns the processes by which cells acquire positional information. There are two general models for how patterns form: use of a morphogen gradient, and sequential induction. The morphogen gradientmodel involves the production and release of a diffusible chemical signal called a morphogen. Morphogen release creates a concentration gradient, with high concentrations of morphogen close to the source, and low concentrations farther away from the source. Exposure to different threshold levels of morphogen leads to different cell fates. In the example below, very high concentrations of the morphogen (above threshold 3) lead to the blue fate, medium levels of morphogen (betweens thresholds 2 and 3) lead to the red fate, and low levels of morphogen (between thresholds 1 and 2) lead to the purple fate. In this way, different amounts of one chemical signal can create a complex pattern.
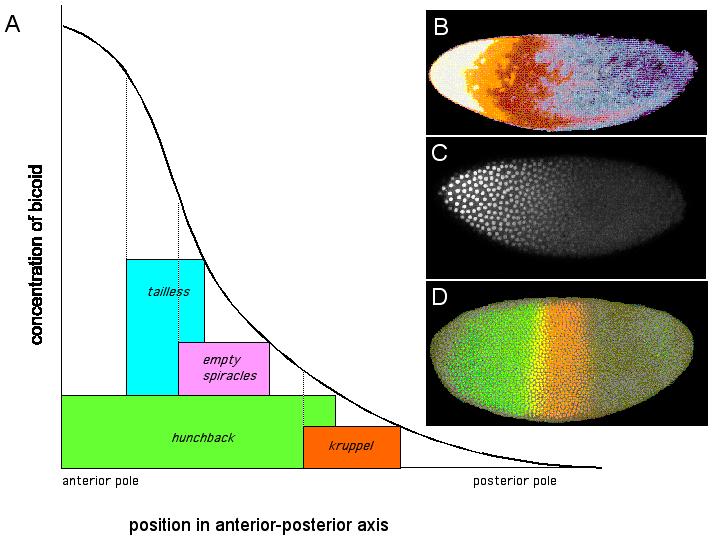
What is an example of the use of a gradient in pattern formation?? The very first step in patterning the embryo of the fruit fly, Drosophila melanogaster, is a good example of pattern formation by a gradient. We'll talk more about Drosophila development next week. But for now, let's just use it as an example of this important concept. Bicoid is a transcription factor which turns on different genes in different levels - acting as a morphogen gradient. In this way, the four genes shown in part A (tailless, empty spiracles, hunchback, and kruppel) are found in different locations within the Drosophila embryo, as a result of the amount of Bicoid protein at a particular location in the embryo. After fertilization, bicoid mRNA from the mother fly begins to be translated into Bicoid protein in the Drosophila zygote. The computer-generated image B shows how the Bicoid protein diffuses through the egg forming a gradient. High concentrations of Bicoid protein are shown in white on the left (anterior) end of the zygote, and low concentrations are shown in blue on the right (posterior) end. Image C shows Bicoid protein in the nuclei of a Drosophila embryo after a number of rounds of mitosis. Notice that the nuclei in the anterior end (left) have more Bicoid protein than those in the posterior end (right) . Image D shows Kruppel protein in orange and Hunchback protein in green. The region where the two proteins overlap is yellow. The colors come from fluorescent dyes attached to antibodies that bind specifically to these proteins. images B and D from Robert
J. Huskey
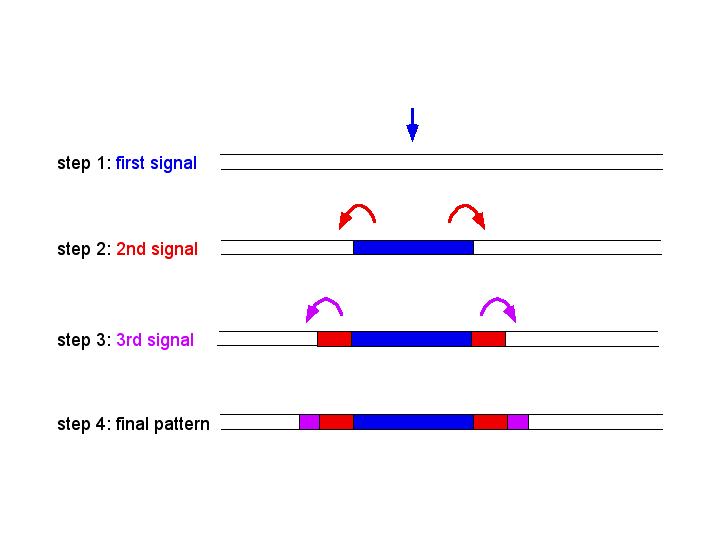
The sequential inductionmodel involves the production and release of a series of chemical signals. Signal 1 leads to the blue fate and production of signal 2. Signal 2 is received by neighbor cells, and leads to the red fate and production of signal 3. Signal 3 is then received by neighbor cells, and leads to the purple fate. In contrast to the morphogen gradient model, multiple chemical signals are required to create the pattern.
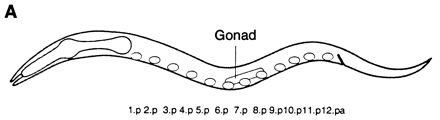
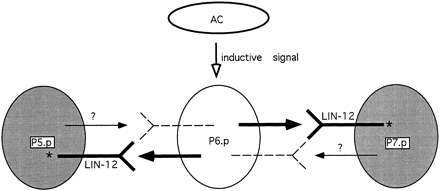
What is an example of the use of sequential induction in pattern formation?? The development of the vulva (a ventral opening used for copulation and egg-laying) in the soil nematode, Caenorhabditis elegans, is a good example of pattern formation by sequential induction.
Thanks to David Marcey for construction of some of the images shown above. |